Ochotona horaceki is a small species of pika of the subgenus Ochotona (theoretically could be considered as), related to the extinct steppe-adapted pikas that during the pleistocene inhabited most of Europe. These European forms are thought to be of the “O. pusilla” species clade, hence, their closest extant relative is unsuprisingly the steppe pika (Ochotona pusilla).
It was found in the western part of the Slovak Karst mountain range, and described in 2004 as a new species based on several unique traits. It lived during the Early Pleistocene (Calabrian), Biharian Small Mammal age.
It is time to take a dive into more very scarcely understood “rodents” of Eocene China, to actually see how they might have looked like when they were still alive.
The monophyly of the cohort Glires, has been disputed for many years. In the present, new genetic studies confirm the monophyly between the two orders, rodents and lagomorphs. Glires however, includes many ancestral, primitive and basal forms that cannot be stricly defined as a rodent or a lagomorph. Glires can be actually better divided to duplicidentata and simplicidentata. Duplicidentates (two tooth) includes the lagomorphs and other closely related forms (with two pair of front incisors) that are not part of the crown group lagomorpha. The same applies for simplicidentata (one tooth), Rodents and their closely allied forms. Inside simplicidentata, another order sister to Rodentia was established to lump some non-rodent simplicidentates called Mixodonta.
One monotypic genus of one of these obscure basal gliroids is Rhombomylus turpanensis. It is known from a lot of different specimens, also from different stages of ontogenic development. Its anatomy is derived to a certain extend, more so, than that of its very close relative Matutinia nitidilus, which possesses more primitive dentition and skull, and it is also smaller in size.
Here is my musculoskeletal reconstruction of the anatomy of Rhombomylus turpanensis. The postrcranial anatomy of this species remains largely obscure. It was likely an unspecialized terrestrial, non-cursorial form with some capabilities for jumping. The hindlimbs are much longer than the forelimbs.
And the life reconstruction:
Here is my another reconstruction of a close relative of Rhombomylus of a different genus, Matutinia nitidilus. I see these animals resembling perhaps the living pikas (Ochotonidae) or the mountain beaver (Aplodontiarufa) and perhaps also the true rats (Rattus). Terrestrial herbivores, living in burrows and dens. They would also supply their diet with animal protein if given the change. These are my guesses of what would be their best modern analogue, althought their trophic ecology did not had to be exactly identical to the before-mentioned examples.
Basal gliroids and their closely related clades remain largely obscure. I hope more research will continue on this group to shed some light on these creatures.
References:
SUYIN TING, JIN MENG, MALCOLM C. McKENNA, and CHUANKUEI LI “The Osteology of Matutinia(Simplicidentata, Mammalia) and Its Relationship to Rhombomylus,” American Museum Novitates2002(3371), 1-33, (21 June 2002). https://doi.org/10.1206/0003-0082(2002)371<0001:TOOMSM>2.0.CO;2
JIN MENG, YAOMING HU, and CHUANKUI LI “THE OSTEOLOGY OF RHOMBOMYLUS (MAMMALIA, GLIRES): IMPLICATIONS FOR PHYLOGENY AND EVOLUTION OF GLIRES,” Bulletin of the American Museum of Natural History 2003(275), 1-247, (28 February 2003). https://doi.org/10.1206/0003-0090(2003)275<0001:TOORMG>2.0.CO;2
I present another short post and reconstruction of another not very well-known prehistoric tooth-digging rodent, of Early Oligocene.
So, here we have it, Tsaganomys altaicus. It belonged to the family Tsaganomyidae, comprising 3 genera and 5 species. Tsaganomys was native to Oligocene and possible early Miocene of Asia. It possesed a protrogomorphous zygomasseteric system (the most primitive type found in rodents) of the skull and a hystricomorphous mandible. It had massively developed almost circular zygomatic arches, large temporal fossa, prominent sagittal and occipital crests, hypsodont (high crowned) dentition and of course, procumbent incisors, among other traits.
The phylogenetic position and ancestry of tsaganomyids appears to be unresolved. Due to several superficial similarities, tsaganomyids were associated with bathyergids, cylindrodonts and hystricognaths. Now it’s clear that this group is not closely related to any of the before mentioned clades. For example, the bones of the middle ear, malleus and incus are not fused as in Bathyergidae, or for example the incisor microstructure is quite different from that of the cylindrodonts. Also, they lack other derived features common for hystricognaths. For now, tsaganomyids shall remain Rodentia incertae sedis.
For more detailed description, I highly suggest reading the original paper I referenced (under the drawing).
Here is my reconstruction of Tsaganomys altaicus. Missing postcranial proportions are based off of Bathyergus and soft tissues are based off of several different species of zokors (Myospalacinae).
Tooth-digging has convergently evolved multiple times among unrelated clades of fossorial rodents throughout the history of the cenozoic. Contrary to the mole-like digging in which the forelimbs act as shovels, tooth-diggers possess unspecialized, relatively small feet, all the digging is done by the one pair of incisors and the head. One clade of these animals is the Spalacinae.
Based on almost identical shape of the mandible, Rhizospalax was once thought to be a spalacid, but detailed examination of its skull and dentition showed that it was of the sciuromorphous, castoroid type. It possessed a large P4, which when present in muroids, is definitely smaller than M1 (spalacids are muroids). The dentition of Rhizospalax was therefore 1.0.1.2/1.0.1.2 (last premolar substituted the first molar – P4, M1, M2) unlike a typical muroid which has only three true molars. Moreover, its molar pattern is less complex as in muroidea, indicating a different ancestry. Another trait that links this taxon to castorids is the shape and position of the infraorbital canal.
Fossorial beavers are known from Oligocene of North America. As a french find, Rhizospalax is a proof that, tooth-digging castorimorphs likely once inhabited the whole holarctic realm, and occupied ecological niches now filled by the still-living Bathyergidae, arvicoline tribe Ellobuisini and Spalacidae.
Here is shown my life reconstruction of Rhizospalax poirrieri. Missing components are based due to ecomorphological similarities, on Spalax, Tachyoryctes and Capacikala.
A new rodent from the Upper Oligocene of France. Bulletin of the AMNH; v. 41, article 18. Gerrit Smith Miller, James Williams Gidley, B Poirrier, Claude Gaillard
Succeeding the Eemian or the Last Interglacial, the Last Glacial period (115, 000 -11, 000 years ago) began. It is a well known fact, that during this time, cave bears (Ursus ex. gr spelaeus) were widespread in Eurasia. This lineage was closely related to recent brown and polar bears. Althought rivaling their extant relatives in terms of dimensions (and on average being heavier), cave bears were specialized herbivores. It can be deduced from numerous factors (isotope analysis, tooth microwear…). There is no doubt they would be a formidable adversary when confronted with our ancestors.
My reconstructions and size chart of several forms of cave bears present in Europe during the Last Glacial.
Cave bears (Ursus ex. gr spelaeus) were very diverse, with different forms living on different localities and having different prefences (altitude and perhaps also plant types..). In central and eastern Europe, Ursus ingressus was one of the largest forms, occupying habitats below the ranges of the smaller, high alpine subspecies such as Ursus spelaeus ladinicus and Ursus spelaeus eremus.
As it has been found, U. ingressus and U. s. eremus were living sympatrically for 15,000 years in Austria, with no apparent interbreeding. This is an evidence, that they must have niche partitioned and were quite different ecologically.
There were also a lot of different forms. For example, the classical Ursus spelaeus spelaeus was found in western Europe.
The brown bears (Ursusarctos) lived alongside cave bears, but were more carnivorous than they are today. This was to avoid competition.
Originally thought to be a wolverine-like animal, (Plesiogulo relative) then viewed as an amphicynodontine ursoid and the third time, we might got it entirely correct… Drassonax harpagops was one of the first definitive ursids (true bear).
Drassonax is a genus with just incomplete remains preserved, like upper maxillary teeth and lower jaws. The only known basal ursoids (ursoids include the Ursidae as well as forms already showing ursid traits but still not belonging to the family) with preserved whole skeletals (therefore giving us much more clues about their anatomy and biology) are Amphicynodon and Eoarctos. Based on dental characteristics, Drassonax appears to be a close relative of the former.
My reconstruction of Drassonaxharpagops, one of the early basal ursids. In life it would resemble a small agile predator like a marten rather than modern bears.
Here is a rough hypothetical sketch of how could the skull of Drassonax look like. The proportions are based off of close relatives such as Amphicynodon and Campylocynodon.
The first true caniforms very likely evolved in North America, as it’s indicated by the oldest and most primitive forms of both arctoids (bear and musteloid ancestors) and cynoids (canids) appearing there as early as in the middle Eocene, and in the case of the first ursoids, the genus Subparictis in the Late Eocene during the Chadronian land mammal age, approximately 38-33 million years ago.
They all started out as largely very active mesocarnivores of rather compact dimensions with adaptations for arboreal or scansorial lifestyle. Drassonax likely possesed great climbing and grasping abilities, also being able to rapidly accelerate. As with its close relative Amphicynodon, it is possible that also Drassonax could reverse its feet, perhaps even up to 180°. This would enable it to descend and ascend trees headfirst.
Perhaps one of the best still-living examples of these small carnivorans that we can compare to our extinct species, in terms of ecology is the enigmatic african palm civet (Nandinia binotata). As well as with others, they resemble the ancestral body plan of the earliest arctoids that all others subsequently evolved from.
Baskin, J. A., and R. H. Tedford. 1996. 24. Small arctoid and feliform carnivorans; pp. 486–497 in D. R. Prothero and R. J. Emry (eds.), The terrestrial Eocene-Oligocene transition in North America, Part II: Common vertebrates of the White River Chronofauna. Cambridge University Press, Cambridge.
My favorite aquatic birds are the grebes (Podicipedidae). When you get the opportunity, it’s always an exciting moment observing in its natural habitat an individual diving and hunting or a family being together.
During the Late Miocene (Tortonian age, roughly 11-7 milion years ago) large part of Eastern Europe was covered by a megalake (a remnant of the inland sea) called the Paratethys. There, among others, numerous species existed that would be superficially recognizable today. These were waterfowl, loons, grebes, cormorants, turtles, seals and even pygmy whales.
One of the birds found in this locality is the grebe Podiceps miocenicus. Judging from the broken humerus preserved, it was similar to the crested grebe (Podiceps cristatus) but somewhat larger in size.
My reconstruction of Podiceps miocenicus, a grebe from the Paratethys megalake of Eastern Europe (Moldova).
The enigmatic and unique molluscivorous carnivoran Kolponomos, has been dubbed by the nickname “the beach bear”. Mostly reconstructed as a true ursid, adapted to live in littoral enviroments, preying on hard-shelled seafood such as molluscs.
But was it really like that? The material of Kolponomos, is well, fragmentary… lacking significant postcranial bones. Althought it is quite likely that it was a littoral molluscivore – judging by the skull and dentition, its ancestry and life appearance remains a bit of a mystery.
Speculative reconstruction of Kolponomos newportensis as a stem-pinniped.
The part of the body we need to focus on is the skull. None of the upper teeth are preserved, but the mandibular are. The large round-shaped teeth are worn out so much so, that the original unworn structure of the cusps (on mostly molars) is not known. This wearage is an evidence for a durophagous lifestyle in a similar manner to the extant sea otter (Enhydra lutris). However, the skull structure of the two is not so similar. Kolponomos had a very pronounced chin that worked like an anchor in aiding to dislodge the clams tightly stucked to the sea floor, by the upper portion of the skull. This is a different strategy, that is not seen in sea otters or in any mammal alive today. In foraging, Kolponomos enhanced mandibular stiffness over mechanical efficiency. The stiffness of the lower jaw is an ursid trait, unlike that of the sea otter which utilizes the mechanical efficiency.
As of now, the phylogenetic position of Kolponomos is uncertain. If it is indeed not a true ursid, it might be a member of the clade Amphicynodontidae. This group contains sort of “intermediate” forms between the terrestrial arctoids and pinnipeds, so called stem-pinnipeds. This does not mean that Kolponomos is ancestral to living seals, but that it represents an offshoot of the numerous stem-pinnipeds that went their own evolutionary path, leaving no descendants in the present day.
Only recently, new light has been shed on the origins of pinnipeds and their relationship to ursoids and musteloids. With a handful of transitional forms being recognized properly. But still, the phylogenetic position of amphicynodontids can vary based on the research. It might be that in future this family will no longer be in use, when all the taxa in it get a proper phylogenetic position (either a stem-pinniped, stem-ursid or a musteloid…)
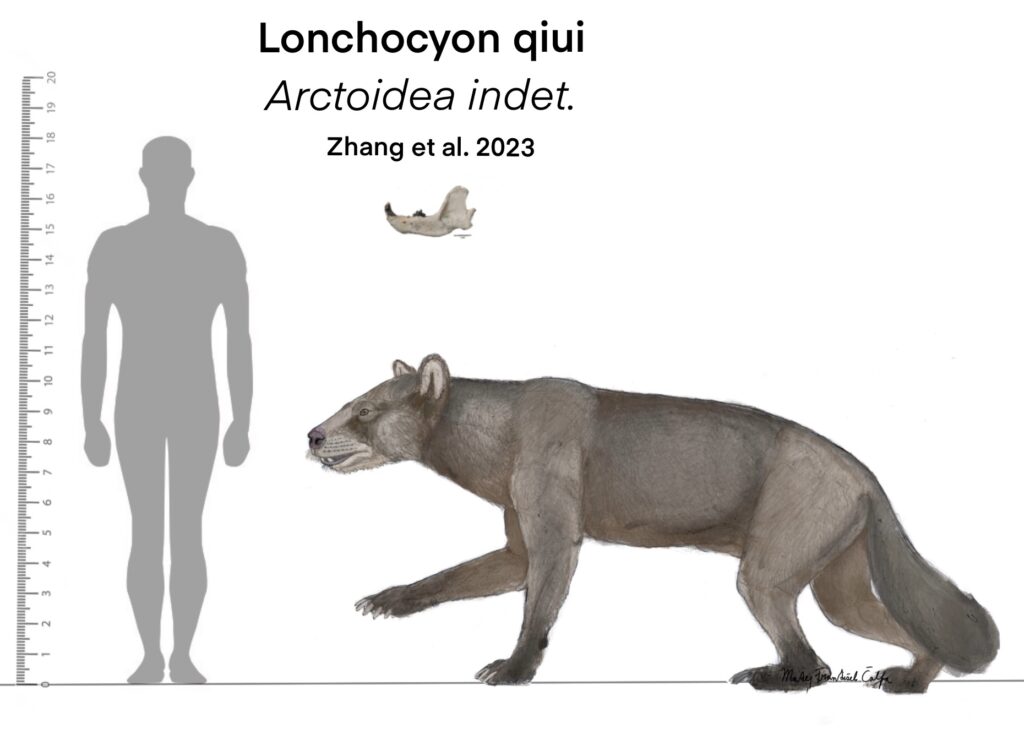
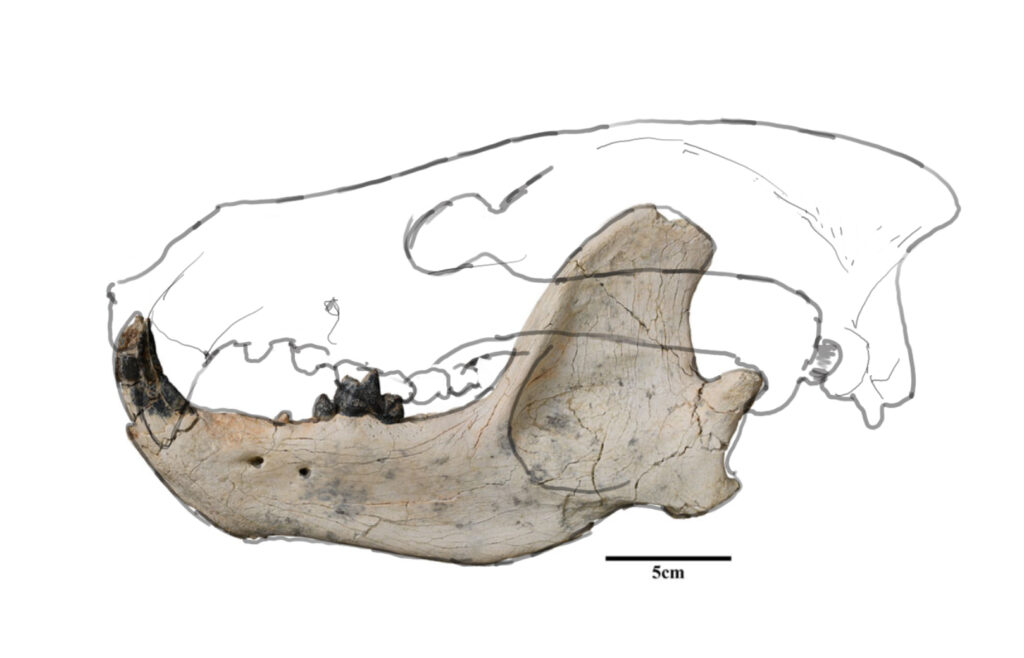
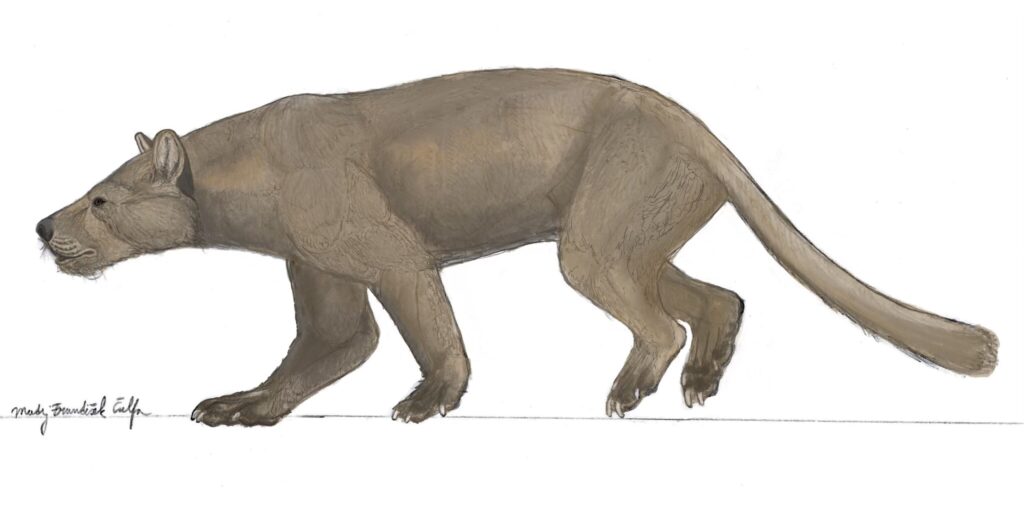
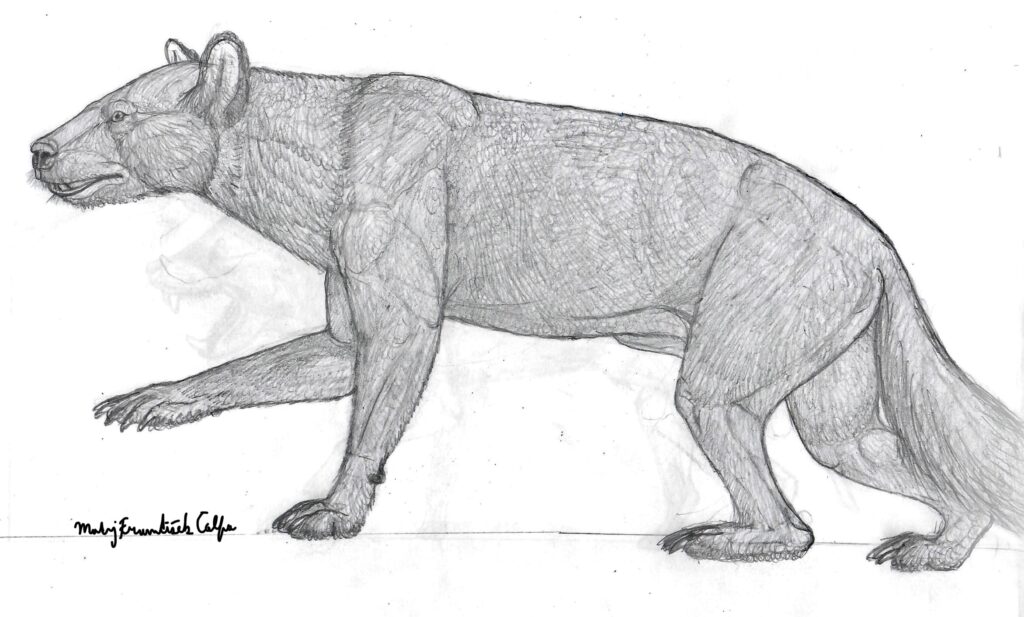
Quite recently, in 2023, a new species of a large carnivoran from the Late Eocene of Mongolia was described. The only material known of Lonchocyon qiui is sadly quite fragmentary, with only one hemimandible, c1 (canine), p4 (4th premolar) and m1 (carnassial) being present.
The skull and postcranium is based off of other hypercarnivorous arctoids, that show large resemblance in the morphology of the mandible and therefore, occupied similar ecological niche.
Based on the lower jaw alone, it becomes clear, that this species was a hypercarnivore. That is, it obtained more than 70% of its calories by meat, either hunted on its own or scavenged on. This is indicated by the robust canine, deep jaw, sectorial trigonid of m1 (that is, specialized for shearing) and strongly reduced premolars. But, as authors of the paper state, there are several hypercarnivorous arctoids that posses functional non-reduced p4 (opposed to our species).
One characteristic of Lonchocyon that could be considered “primitive” or unspecialized, is the retention of unreduced post-carnassial teeth (m2, m3) which usually do the crushing job in carnivorans. In other carnivoran hypercarnivores, these teeth tend to be strongly reduced.
Taking it all to consideration, L. qiui, could consume both flesh and hard objects like bone.
Here, a hypothetical skull is shown:
Right now, the question remains: Was Lonchocyon an amphicyonid or a true bear of the subfamily hemicyoninae? It’s not known yet. What is known however, is, that it represents an early offshoot of one of these groups, appearing quite early in their history (Late Eocene).
During the cenozoic, the suborder Caniformia has produced several lineages of carnivores, which cannot be compared to any living caniforms, hence, occupied a niche that either today remains vacant or is taken by a non-caniform. Such example would be the amphicyonids (bear-dogs) or the ursids (true bears) of the subfamily hemicyoninae. Contradictory to the name “bear-dog”, later, more specialized amphicyonids (amphicyoninae, thaumastocyoninae..) show a combination of skeletal traits and articulations rather resembling a mix between a bear and a big cat. Having a tendation to become hypercarnivorous, with the ability to deliver powerful, deep canine bite, combined with well developed pronation-supination movement, one would imagine these predators as solitary, ambush hunters and scavengers, much more like big cats and bears than any canid or hyaena. One representative of these robust carnivores is Ammitocyon:
Does this mean that this is exactly how we can imagine Lonchocyon to look like? Perhaps yes. Until more of its material is found and described it’s hard to tell with absolute certainty how exactly L. qiui was built.
This fascinating caniform remain to remind us of our humble knowledge of the history and evolution of the Caniformia clade, that in the past produced one of the most fearsome mammalian land predators that roamed the whole Afroeurasia and North America.
A medium dog-sized carnivore that is able to take down on its own ungulates, survives the harshest winters far north and does not hibernate..
Well, there is an actual animal like that, the wolverine (Gulo gulo).
Wild wolverine in Finland. Image: pixabay
The wolverine is a great example of an opportunistic predator and scavenger. It will feast on anything it stubles on, taking huge advantage of any opportunity given.. While most of the time it’s looking for smaller prey and carrion, it can take down ungulates much larger than itself such as reindeer. It’s also a scavenger, with a very acute sense of smell, capable of detecting carcasses covered up by snow with great accuracy.
With frost and water resistant fur and paws of very large size, that help them to stay on top of deep snow, its anatomy is greatly adjusted to the conditions of arctic and boreal forests.
Its lifestyle and habits are reflected in the skull and dentition. When compared to their closest relatives, it posseses enormously enlarged carnassials (P4 and m1) to crush bones and also, the upper molars are rotated in 90° angle which enables them to tear off frozen solid meat. Overall, the skull looks like that of an absolutely formidable predator.
The wolverine is a member of Mustelidae, that means that it’s basically a giant marten or a weasel rather than a small bear. The members of the weasel family are known to have absolutely enormous capabilities relatively for their body size, which is only confirmed in this species.
To look a little bit at its background and history, among mustelids, the wolverine is most closely related to martens (Martes) and fisher (Pekania). Together forming the subfamily Guloninae. To whose of these is the wolverine more closely related isn’t 100% sure. It’s likely that their common ancestor looked similar to the fisher Pekania occulta.
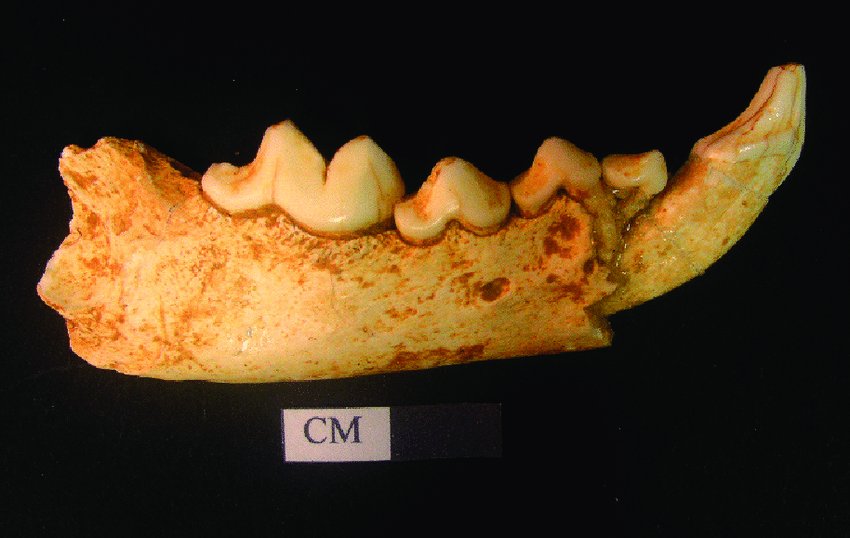
I had the idea of reconstructing some of its ancestors, or closely related extinct species. What came to my interest, is this fragment of the lower jaw of the ancestor of wolverine, being classified in its own species Gulo schlosseri, from Żabia cave in Poland.
To highlight the proportions and life appearance of this animal with the best possible accuracy, I combined this material with skeleton of modern wolverine, which gave me a good base for my drawing.
With a decent “strolling” pose and proportions, the muscles were added very roughly. I chose it yawning, to show its stunning hypercarnivorous dentition mentioned above.
Since the animal, in life would be all covered in fur, no muscular anatomy would be visible. This has made the job a bit easier. Here’s the outline of it.
Then, the details…
And finally, coloration. Althought color variations in individual wolverines can be suprisingly variable, for this one, I chose more darkly-toned individual. Perhaps, all these populations were darkly colored because of their shared ancestry with the fishers (Pekania)? This is a matter of speculation :).
Here we have it. The reconstruction of Gulo schlosseri, an ancestral wolverine from Early Pleistocene. Acting more or less like its living descendants, it surely would have been an amazing experience encountering one of these in their natural habitat.