This article is a synthesis of referenced literature on the topic, and apart from my life reconstructions I have not conducted any original research on the group. I wrote it in a way that it may resemble an actual scientific article a little, so it helps to highlight and organize information in a better manner. If you are aware of any paper I have not referenced, please contact me, so I can make this list even more comprehensive.
Anomaluroidea is a poorly studied superfamily of elusive rodents of mainland African origin and present zoogeographical distribution. This study presents a synthesis of all dental and skeletal remains attributed to Anomaluroidea throughout the tertiary, with an introduction to the ecomorphology, evolution and systematics or the group. In total 216 specimens in 5 families are included in the list.
Introduction to the group’s taxonomy, phylogeny and morphology:
The superfamily Anomaluroidea contains in total of 7 extant species distributed in 2 families. Anomaluridae includes the subfamilies Anomalurinae and Idiurinae. Anomalurinae includes 4 species of scaly-tailed squirrels, the Lord Derby’s scaly-tailed squirrel (Anomalurus derbianus), Pel’s scaly-tailed squirrel (Anomalurus pelii), dwarf scaly-tailed squirrel (Anomalurus pusillus) and Beecroft’s flying squirrel (Anomalurus beecrofti) which is the most divergent of the anomalurines and is sometimes included in its own genus Anumalurops (Fabre et al. 2018). Idiurinae includes the long-eared flying mouse (Idiurus macrotis) and the pygmy scaly-tailed flying squirrel (Idiurus zenkeri) (Schunke et Hutterer 2017). The family Zenkerellidae includes only a single species, the Cameroon scaly-tail (Zenkerella insignis), which has not been directly observed by a western mammalogist until 2017 (Dinets, 2017).Together with the springhares (Pedetidae), they form the suborder Anomaluromorpha, one of the four main suborders of rodents (Fabre et al. 2018). Based on palaeontological and molecular evidence, Anomaluroidea is a remarkably ancient and conservative clade, with the divergence time of the crown group Anomaluroidea being estimated during the early or middle Eocene (Heritage et al. 2016). Originally, the Zegdoumyidae, recorded from the Middle Eocene, was thought to be the basalmost anomaluroid family (Marivaux et al. 2014). There is some uncertainty regarding their anomaluroid affinities, however (Coster et al. 2015). Later, using the Bayesian tip-dating analysis, the springhare (Pedetidae), an anomaluromorph but not an anomaluroid (scaly-tailed squirrel) was found to be more closely related to Anomaluroidea than the Zegdoumyidae, therefore placing Zegdoumyidae as stem Anomaluromorpha (Seiffert et al. 2024). This then excludes them from the list.


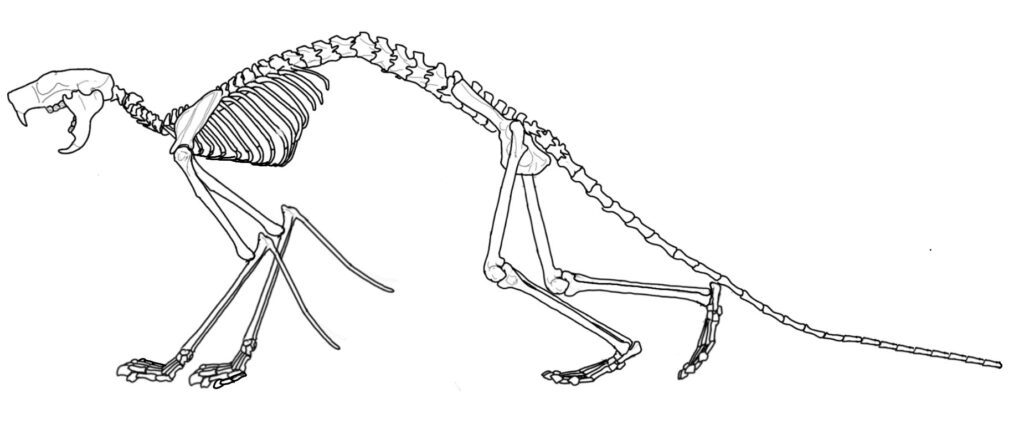
Extant anomalurids posses several unorthodox gliding adaptations not recorded in any other living volant mammal (colugo, pteromyin squirrels and sugar gliders) Among these, a specialized rod-like cartilage on the arm, originating from the olecranon proccess of the proximal ulna (Amador, 2022), and peculiar scales on the ventral side of the proximal part of the tail. (Panyutina et al. 2020). Furthermore, anomaluromorphs (Anomaluroidea and Pedetidae) possess the orbital portion of the infraorbital part of the zygomaticomandibularis as well as the rostral portion, hence exhibiting double origin of the zygomaticomandibularis. The relevance of this trait is in the fact that the orbital portion of the zygomaticomandibularis was thought to be absent in rodents (Druzinsky et al. 2011), and while it has been eventually confirmed in Bathyergidae (Cox & Faulkes, 2014; Cox et al., 2020), the later is not of double origin as in the former. Therefore, this structure of the infraorbital part of the zygomaticomandibularis is unique to Anomaluromorpha (Da Cunha et al. 2021).
Materials and methods:
The data in the list were gathered from published literature spanning the period 1973-2024. Included were papers, books, dissertations, abstracts, and posters, for a total of 24 publications. They were found using Google Scholar or searching for author’s publications online. Any possible unpublished, privately-owned or undescribed specimens were excluded. Individual specimens and localities are not cited separately if the species was published in a single paper. In such case, the authors are cited solely at the begging of the list of a given species. The life reconstructions were done traditionally, using paper and a mechanical pencil, and then colored digitally. With the smaller species (Shazurus, Argouburus), allometric growth was considered, which directly affects the body proportions – head to body size ratio. The cranial and postcranial proportions of each species were reconstructed based on its closest living relative, i.e. Zenkerella, Anomalurus, or Idiurus. Original specimens were digitally scaled to the nearest 0.1 mm.
Results:
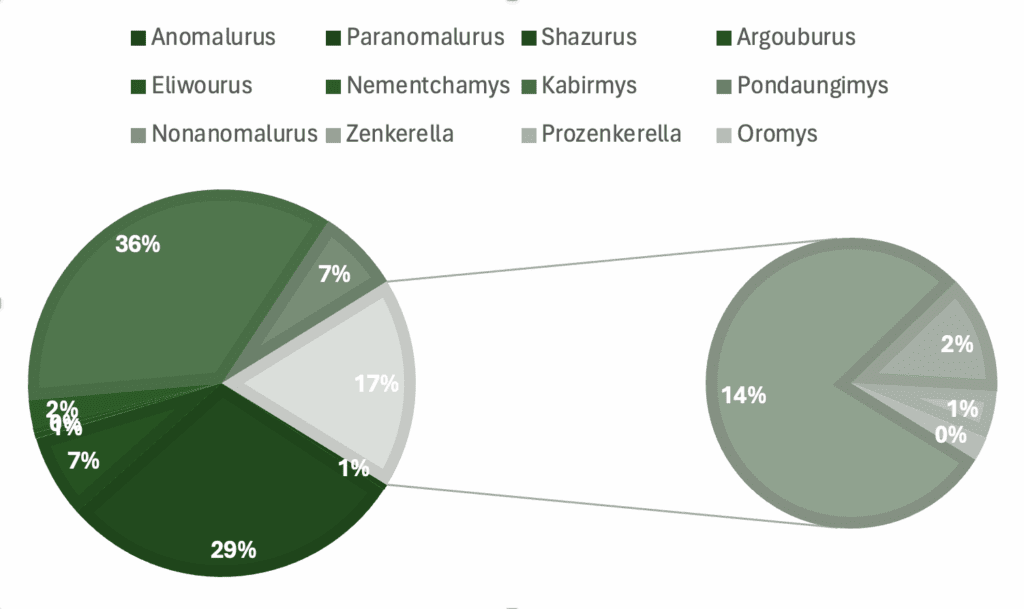
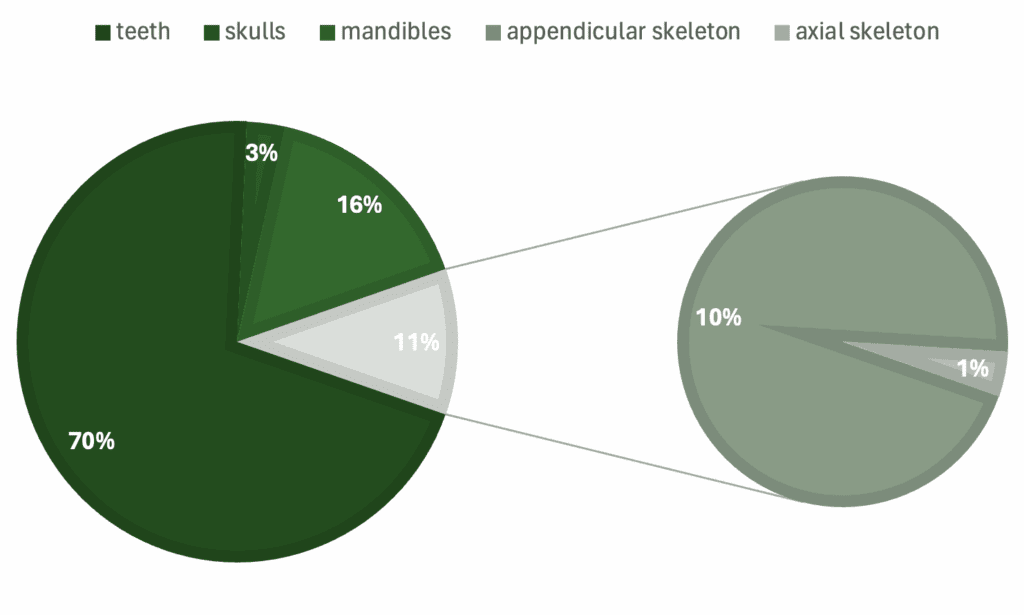
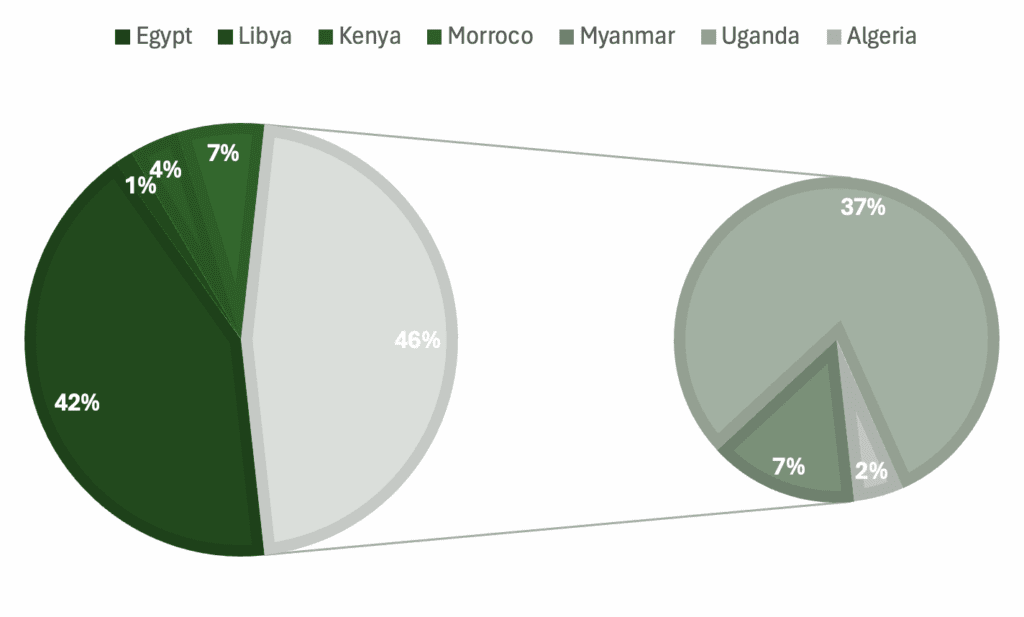
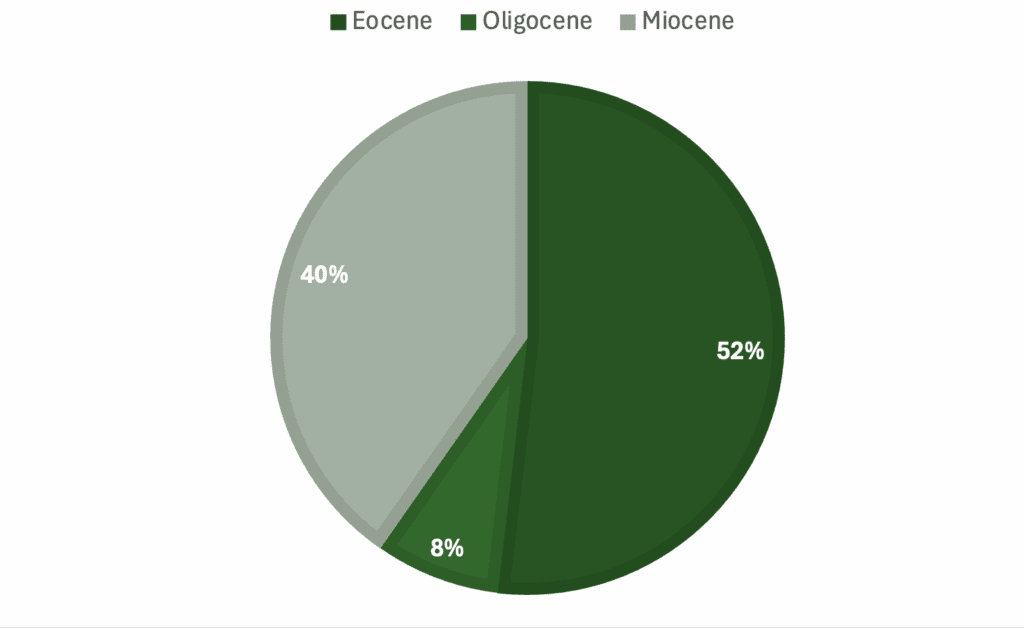
Graphs showing percentage of specimens regarding a) genera, b) skeletal material, c) country of origin, d) epoch.
List:
Anomaluridae
Family that includes all the extant gliding anomaluroids, Anomalurus and Idiurus. Members of this group possess a rod-like structure originating from the proximal ulna that enchances gliding locomotion, it is a unique structure only confined to this clade. Some of the species included in this family in the list are not explicitly described in cited literature as belonging to Anomaluridae sensu strictio, but are very frequently compared to the latter based on the profound dental similarities. That is why they are listed under this family.
Anomalurus (Waterhouse, 1843) – middle Miocene, Kipsaraman, Muruyur Beds, Kenya; in total, 1 fossil specimen listed.
(Winkler et al. 2010)
KNM-TH 19439 – left M3,
Paranomalurus bishopi (Lavocat, 1973) –Early Miocene (approximately 20 Ma); Napak IV, V, XV, XXX, XXXI, Northeastern Uganda; the largest species of Paranomalurus, volant, generally thought of as phenotypically very similar to the extant Anomalurus; in total, 43 specimens listed.
(unable to provide description of the type specimen, see (Lavocat, 1973))
(Winkler et al. 2010)
BUMP 82 – neurocranium, right p4-m3
(Bento da Costa; 2020) – no description to individual specimens provided
Teeth – NAP IV 76’09; NAP IV 5’12; NAP IV 165’12; NAP V 9’04; NAP V 191’08; NAP V 246’11; NAP V 146’13; NAP V 7’15; NAP V 49’16; NAP V 83’16; NAP XV 97’09; NAP XV 142’09; NAP XV 155’09; NAP XV 57’14; NAP XV 25’16; NAP XXX 196’14
Mandibles – NAP I 308; NAP I 42’99; NAP IV 50’06; NAP IV 58’08; NAP IV 78’10; NAP IV185’12; NAP IVlower 210’12; NAP IV 25’18; NAP IV 95’18; NAP V 65S; NAP V 24’02; NAP V 36’06; NAP V 25’10; NAP V 98’15; NAP IX AUG 62; NAP XV 171’07; NAP XV 172’07; NAP XV 165’08; NAP XV 53’09; NAP XV 43’10; NAP XV 33’14; NAP XV 16’15; NAP XV 73’16
Ulnae – NAP IV 185’11; NAP V 51’03; NAP XV 64’12
NAP XV 113’07 – right distal humerus

Paranomalurus walkeri (Lavocat, 1973) – Early Miocene (approximately 20 Ma); Napak IV, V, XV, XV, XV, XXXI, Northeastern Uganda; dentition similar to P. bishopi but smaller in size, volant, generally thought of as phenotypically very similar to the extant Anomalurus; in total 13 specimens listed.
(unable to provide description of the type specimen, see (Lavocat, 1973))
(Bento da Costa; 2020) – no individual description to specimens provided
Teeth – NAP IV 64 428; NAP IV 108’13; NAP IV 68’14; NAP V 145’13; NAP V 172’15; NAP XV 146’09; NAP XV 6’12; NAP XV 36’18; NAP XXXI 56’16
Mandibles – NAP XV 146’09; NAP XV 6’12
Femora – NAP XV 375’08; NAP XV 99’09
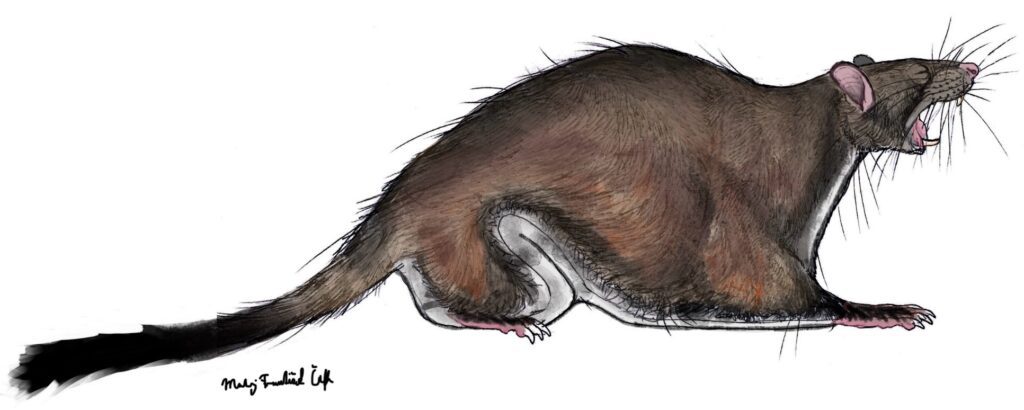
Shazurus minutus (Sallam et al. 2010) – Earliest Late Eocene (approximately 37 Ma); Birket Qarun Locality 2 in the Fayum Depression of northern Egypt; a miniscule species of an anomalurine rodent, with a dentition similar to Paranomalurus; in total, 15 specimens listed.
DPC 23305A – left p4; DPC 21294A – right p4; DPC 21747A – right p4; DPC 21488B – left m1; DPC 21220B – right m1; DPC 21293L – left m1; DPC 21296A – right m2; DPC 21500B – right m2; DPC 21358A – left m2; DPC 22442B – left m2; DPC 23305B – right P4; CGM 83701 – left M1; DPC 21538A – right M1; DPC 21221A – right M2; DPC 21452A – left M2
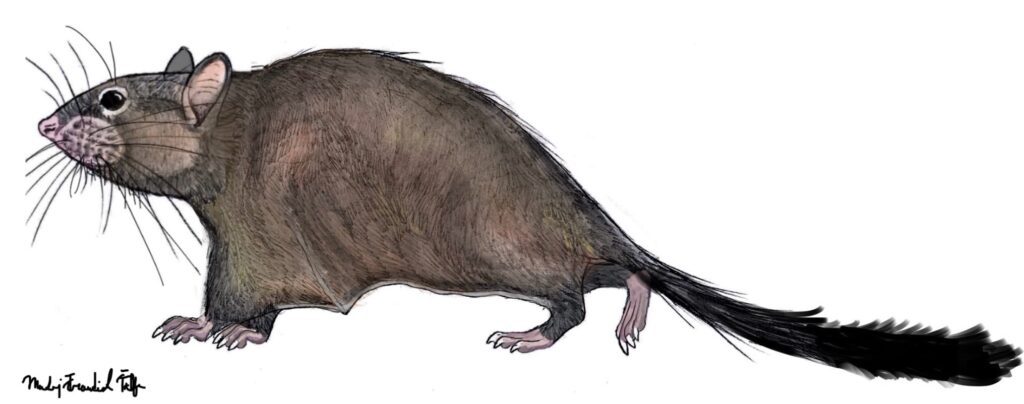
Argouburus minutus (Marivaux et al. 2017) – Earliest Oligocene (approximately 33 Ma); Upper Samlat Formation, westernmost part of the Sahara, Morroco; a miniscule species, differs from both Shazurus and Paranomalurus in lacking the Y-shaped structure of the third and fourth transverse crests; in total, 1 specimen listed.
DAK-Arg-010 – Left M3; holotype

Zenkerellidae
Non-volant, scansorial family, including the sole extant species, the Cameroon scaly-tail (Zenkerella insignis). The behavioral ecology of this species is poorly understood (Heritage et al. 2016).
Zenkerella wintoni (Lavocat, 1973) – late Early Miocene (approximately 17 Ma); Moroto II, Kogole, Songhor, Kenya; smaller than Z. insignis; In total, 5 specimen listed.
mandible – (unable to provide more detailed description of the specimen, see (Lavocat, 1973))
(Pickfort et. Mein 2006)
Mor II, 14’98 – left mandible fragment with m1 and m2; Mor II, 33’03 – right molar fragment (left m3?); Mor II, 46’04 – left P4; Mor II, 47’04 – left p4

Prozenkerella saharaensis (Coster et al. 2015) – Early Oligocene; Zallah 7 Incision locality, Continental and Transitional Marine Deposits (CTMD), central Libya; a species very similar to Zenkerella wintoni and Zenkerella insignis, larger than Z. wintoni but slightly smaller than Z. insignis; in total, 2 specimens listed.
Z7I-12 – right m, holotype; Z7I-13 – right M

Oromys zenkerellinopsis (Marivaux et al. 2017) – Earliest Oligocene (approximately 33 Ma); Upper Samlat Formation, westernmost part of the Sahara, Morroco; miniscule species with brachydont and quadrangular teeth, differs from other Zenkerillidae in showing a less-rounded crown outline; in total, 1 specimen listed.
DAK-Arg-009 – left M1 or M2, holotype

Nonanomaluridae
A monotypic family including only the genus Nonanomalurus. The family exhibits postcranial anatomy lacking the typical distal elongation of the appendicular skeleton of the gliding forms such as Anomalurus, being the result of an arboreally and partially terrestrially adapted mode of life without gliding (Pickford et al. 2013).
Nonanomalurus soniae (Pickford et al. 2013) – Uganda Early Miocene (approximately 20.5-18.5 Ma); Napak IV, V, XV, XXX, XXXI, Akisim remnant of the Napak Volcano, Northeastern Uganda; non-volant arboreally adapted species retaining several plesiomorphic characters shared with Pedetidae. Postcranial proportions drastically different from contemporaneous anomalurids, possibly indicating a lifestyle similar to extant galagid primates; in total, 25 specimens listed.
(Lavocat, 1973)
KNM SO 788 – left maxilla P4-M2; holotype
(Pickford et al. 2013)
Nap XV 170’07 – partial skeleton: skull, mandibles, the vertebral column, rib cage, both scapulae, complete left humerus and radius and parts of the ulna, broken right humerus, ulna and radius and disarticulated wrist and manual bones (which are close together) a damaged pelvis, complete left femur and tibia, broken right femur and tibia, talus, ankle and pedal bones. Inside the thoracic cage, four partially ossified long bone diaphyses (two femur and two tibiae) an isolated frontal bone, and an incompletely formed otic bulla, which the authors interpret as the remains of a fœtus (less likely a new born infant).
(Bento da Costa; 2020) – no individual description to specimens provided
teeth – NAP I 58 308; NAP IV 52’07; NAP IV 60’09; NAP IV 14’1; NAP V 8’04; NAP V 140’11; NAP V 245’11; NAP V 83’12; NAP V 155’14; NAP XV 36’11;
postcranial material – NAP I 135’99; NAP V 20’98; NAP V 6’01; NAP V 49’10; NAP V 83’12; NAP XIII 8’14; NAP XV 201’08; NAP XV 59’16; NAP XXX 23’15; NAP XXXI 50’14; NAP XXXI 58’18; NAP XXXII 6’16; NAP XXXII 25’16

Nonanomalurus parvus (Marivaux et al. 2017) – Earliest Oligocene (approximately 33 Ma); Upper Samlat Formation, westernmost part of the Sahara (Morocco); half the dental size of N. soniae, differs from itin the morphology of the upper molars for instance having lower, shorter and thinner transverse crests; in total, 5 specimens listed.
DAK-Arg-011 – left M1, holotype; DAK-Arg-007 – right M2; DAK-Pto-005 – right P4, lingually broken; DAK-Arg-003 – right m2; DAK-Arg-004 – left p4
Nementchamyidae
The basalmost family included in Anomaluroidea. The family exhibits a generalized postcranial morphology, indicating an adaptation to a non-volant lifestyle (Coster et al. 2015; Coster et al. 2019).
Kabirmys qarunensis (Sallam et al. 2010) – Earliest Late Eocene (approximately 37 Ma); Birket Qarun Formation, Umm Rigl Member, Egypt; the largest known anomaluroid, lacking extensive enamel crenulation on the upper and lower teeth, non-volant; in total, 76 specimens listed.
CGM 83698 – right mandible with m1–2, holotype; CGM 08-289A – fragment of left mandible with dp4 and incisor; CGM 21839G – left mandibular fragment with m1; DPC 24494 – fragment of left maxilla with P4 and M1 alveoli; DPC 23305I – right mandible with m1 and alveoli for p4, m2, and m3; DPC 21839F – left M1 or M2; DPC 23305F – left M1 or M2; DPC 21502C – left M1 or M2; DPC 21538F – left M1 or M2; DPC 21221E – left M1 or M2; DPC 21296H – left M1 or M2; DPC 21452F – left M1 or M2; DPC 21500M – left M1 or M2; DPC 21293O – right M3; DPC 21221F – right M3; CGM 08-328B – left M3; DPC 21358G – left M3; DPC 21488G – left M3; DPC 23305F – left M1 or M2; DPC 21374A – left dp4; DPC 21452B – left dp4; DPC 21221B – left dp4; DPC 21360A – left dp4; DPC 21839A – left dp4; DPC 23305C – left dp4; DPC 21294B – right dp4; DPC 21538B – right dp4; DPC 21293M – right dp4; DPC 21358B- right dp4; DPC 21296E – left dp4 or p4; DPC 21500K – right p4; DPC 22442H – left p4; DPC 21839B – left p4; DPC 23305D – right p4; DPC 21538C – left m1; DPC 21221C – left m1; DPC 21294C – left m1; DPC 21358C – right m1; DPC 21360B – right m1; DPC 21374B – right m2; DPC 21452C – right m2; DPC 21500L – right m2; DPC 21538D – right m2; DPC 21839C – right m2; DPC 22442I – right m2; DPC 21360C – right m2; DPC 21221D – right m2; DPC 21296G – left m2; DPC 21294D – left m2; DPC 21306D – left m1 or m2; DPC 21374C – right m3; DPC 23305E – right m3; DPC 21452D – left m3; DPC 21358D – left m3; DPC 21839E – left dP4; DPC 21538E – left dP4; CGM 08-328A – right dP4; DPC 21220D – right dP4; DPC 21360E – right dP4; DPC 21294E – right P4; DPC 21371I – left P4; DPC 21488D – left P4; DPC 21452E – right M1 or M2; DPC 21347D – right M1 or M2; DPC 21747I – right M1 or M2; DPC 22442J – right M1 or M2; DPC 21220E – right M1 or M2; DPC 21358E – right M1 or M2; DPC 21458E – right M1 or M2; DPC 21374E – left M1 or M2; DPC 21371J – left M1 or M2; DPC 21293N – left M1 or M2; DPC 21294F – left M1 or M2; DPC 21358H – left M1 or M2; DPC 21360F – left M1 or M2; DPC 21488F – left M1 or M2

Kabirmys prius (Coster et al. 2015) – Late Middle Eocene (approximately 37 Ma); DT-Loc. 3, New Idam Unit of the Dur At-Talah escarpment, south- central Libya; approximately the same size as K. qarunensis but more plesiomorphic, species with brachyodont and pentalophodont upper molar; in total, 1 specimen listed.
DT3-1 – right M2; holotype
Nementchamys lavocati (Jaeger et al. 1985) – Late Eocene; Nementcha locality, southern slope of Nementcha mountains, Eastern Algeria; species of large size, with dentition typical of the family, strong anterolophid on lower molars, molarized P4, occlusal surface complex, with numerous enamel crenulations; in total, 5 specimens listed.
UON84-25 – right m1; holotype; UON84-26 – left p4/dp4; UON84-27 – right m3; UON84-28 – right P4; UON84-29 – left M3
Pondaungimys anomaluropsis (Dawson et al. 2003) – Late Middle Eocene (approximately 40Ma); Paukkaung locus no. 2 (PK2) and Saba Pondaung (PK4 – Spg), Pondaung area, Central Myanmar (Coster et al. 2019), Bhl locality, Bahin area, Central Myanmar (Dawson et al. 2003); the solely recorded anomaluroid from Asia, the dentition similar to other nementchamyids, non-volant; in total, 15 specimens listed.
(Dawson et al. 2003)
NMMP-KU 0213 – left mandibular fragment m1-3; holotype; NMMP-KU 0048 – right maxillary fragment with P3–4; NMMP-KU 0047 – left m2; NMMP-KU 0049 – left mandibular fragment with m2–3
(Marivaux et al. 2005)
Pkg 157 – partial skull with well-preserved left and right maxillae with P4–M3; Pkg 192 – right DP4; Pkg 193 – left P4; Pkg 194 – right m3; Pkg 195 – left p4; Pkg 196 – left m1; Pkg 197 – left m3; lingual and posterior borders broken; Spg 14 – right p4; Spg 14 – right m1; Spg 14 – right mandibular fragment with p4 and m1
(Coster et al. 2019)
MFP 2014-1 – left astragalus
Discussion and conclusion
The evolutionary history of Anomaluroidea reflects a remarkably ancient and conservative clade (Heritage et al. 2016), likely to be the oldest rodent clade from Africa, that has (to our current knowledge) diversified morphologically very little. Anomaluroidea might potentially represent a rare case of African autochtonous rodents dispersing to Asia as early as Middle Eocene (Coster et al. 2019), which is opposed to most rodent clades that dispersed vice versa, that is, from Asia to Africa, such as phiomorph rodents, which appear to be much more common in fossil localities with anomaluroids present (Marivaux, 2017). This hypothesis is in need of better clarification since there is still the possibility that anomaluroids originated in Asia and dispersed to Africa, with the genus Pondaugimys of Middle Eocene Myanmar representing the oldest undoubted occurrence of this clade, and the only described anomaluroid of Asia. If an asiatic origin of the Anomaluroidea is indeed correct, Zegdoumyidae might not be related to Anomaluroidea at all (Coster et al. 2015). The state of origin for Anomaluroidea shall remain open till more exhaustive material is known to science for better clarification.
The members of the family Nementchamyidae appear to be considerably large in dimension for anomaluroid standards (Jaeger et al. 1985; Sallam et al. 2010), and most likely did not have any adaptation for gliding (Sallam et al. 2010). The same applies for the monotypic Nonanomaluridae, of which the entire skeleton has been preserved, shedding light on its ecomorphology. Nonanomalurus was most likely predominantly arboreal, non-glissant form with partially limited terrestrial lifestyle as well (Pickford et al. 2013; Bento da Costa, 2020). Overall, anomaluroids are known to Eocene sites of North Africa while completely lacking in any Oligocene sites of the same region. The sympatrically known phiomorph rodents do not show this pattern of rehearsal, likely suggesting that a profound alteration of habitats was present in the region somewhere at the Eocene – Oligocene boundary (Sallam et al. 2010)
In conclusion, Anomaluroidea remains as one of the least known clades of extant mammals in the world, and the same applies for the fossil record and evolutionary history. Current picture of the natural history of anomaluroids is partial and incomplete. The aim of this study is to synthesize current record of the clade to acquire a more accurate undestanding of the present knowledge of this group. Until more fossil material is unearthed and empirical studies are done on the topic, not much more can be said about the ecomorphology, systematics, and behaviour of anomaluroids.
References: