My favorite aquatic birds are the grebes (Podicipedidae). When you get the opportunity, it’s always an exciting moment observing in its natural habitat an individual diving and hunting or a family being together.
During the Late Miocene (Tortonian age, roughly 11-7 milion years ago) large part of Eastern Europe was covered by a megalake (a remnant of the inland sea) called the Paratethys. There, among others, numerous species existed that would be superficially recognizable today. These were waterfowl, loons, grebes, cormorants, turtles, seals and even pygmy whales.
One of the birds found in this locality is the grebe Podiceps miocenicus. Judging from the broken humerus preserved, it was similar to the crested grebe (Podiceps cristatus) but somewhat larger in size.
My reconstruction of Podiceps miocenicus, a grebe from the Paratethys megalake of Eastern Europe (Moldova).
The enigmatic and unique molluscivorous carnivoran Kolponomos, has been dubbed by the nickname “the beach bear”. Mostly reconstructed as a true ursid, adapted to live in littoral enviroments, preying on hard-shelled seafood such as molluscs.
But was it really like that? The material of Kolponomos, is well, fragmentary… lacking significant postcranial bones. Althought it is quite likely that it was a littoral molluscivore – judging by the skull and dentition, its ancestry and life appearance remains a bit of a mystery.
Speculative reconstruction of Kolponomos newportensis as a stem-pinniped.
The part of the body we need to focus on is the skull. None of the upper teeth are preserved, but the mandibular are. The large round-shaped teeth are worn out so much so, that the original unworn structure of the cusps (on mostly molars) is not known. This wearage is an evidence for a durophagous lifestyle in a similar manner to the extant sea otter (Enhydra lutris). However, the skull structure of the two is not so similar. Kolponomos had a very pronounced chin that worked like an anchor in aiding to dislodge the clams tightly stucked to the sea floor, by the upper portion of the skull. This is a different strategy, that is not seen in sea otters or in any mammal alive today. In foraging, Kolponomos enhanced mandibular stiffness over mechanical efficiency. The stiffness of the lower jaw is an ursid trait, unlike that of the sea otter which utilizes the mechanical efficiency.
As of now, the phylogenetic position of Kolponomos is uncertain. If it is indeed not a true ursid, it might be a member of the clade Amphicynodontidae. This group contains sort of “intermediate” forms between the terrestrial arctoids and pinnipeds, so called stem-pinnipeds. This does not mean that Kolponomos is ancestral to living seals, but that it represents an offshoot of the numerous stem-pinnipeds that went their own evolutionary path, leaving no descendants in the present day.
Only recently, new light has been shed on the origins of pinnipeds and their relationship to ursoids and musteloids. With a handful of transitional forms being recognized properly. But still, the phylogenetic position of amphicynodontids can vary based on the research. It might be that in future this family will no longer be in use, when all the taxa in it get a proper phylogenetic position (either a stem-pinniped, stem-ursid or a musteloid…)
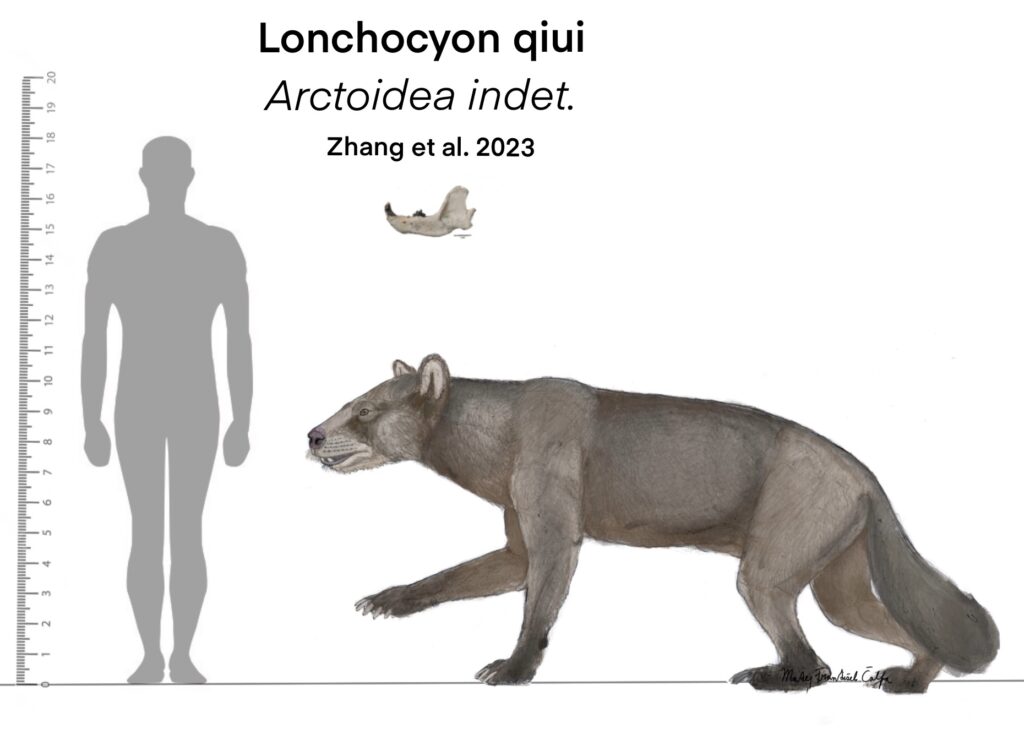
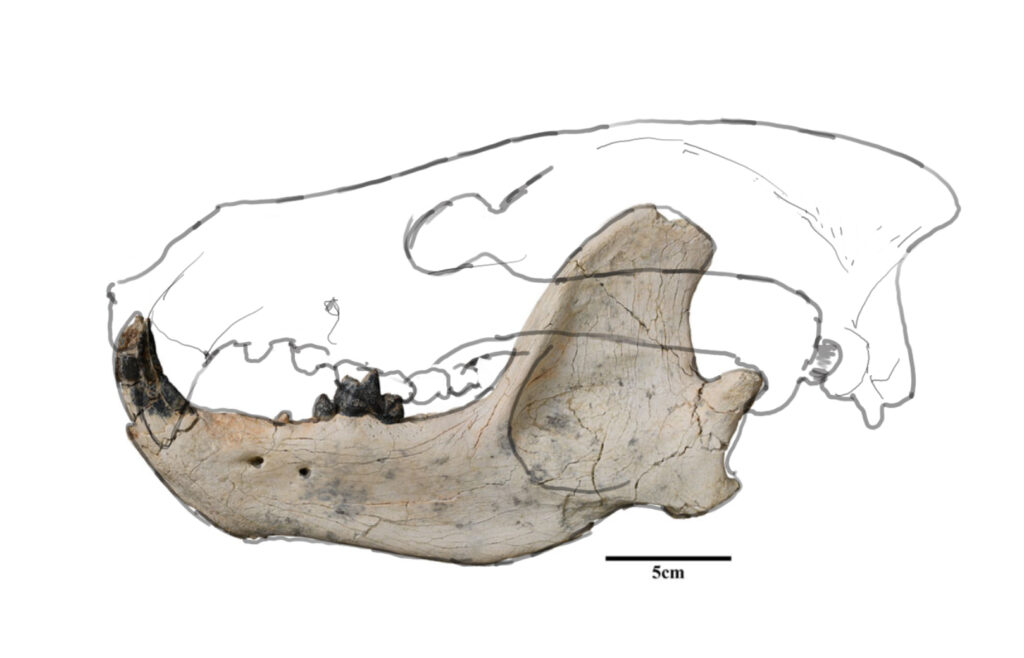
Quite recently, in 2023, a new species of a large carnivoran from the Late Eocene of Mongolia was described. The only material known of Lonchocyon qiui is sadly quite fragmentary, with only one hemimandible, c1 (canine), p4 (4th premolar) and m1 (carnassial) being present.
The skull and postcranium is based off of other hypercarnivorous arctoids, that show large resemblance in the morphology of the mandible and therefore, occupied similar ecological niche.
Based on the lower jaw alone, it becomes clear, that this species was a hypercarnivore. That is, it obtained more than 70% of its calories by meat, either hunted on its own or scavenged on. This is indicated by the robust canine, deep jaw, sectorial trigonid of m1 (that is, specialized for shearing) and strongly reduced premolars. But, as authors of the paper state, there are several hypercarnivorous arctoids that posses functional non-reduced p4 (opposed to our species).
One characteristic of Lonchocyon that could be considered “primitive” or unspecialized, is the retention of unreduced post-carnassial teeth (m2, m3) which usually do the crushing job in carnivorans. In other carnivoran hypercarnivores, these teeth tend to be strongly reduced.
Taking it all to consideration, L. qiui, could consume both flesh and hard objects like bone.
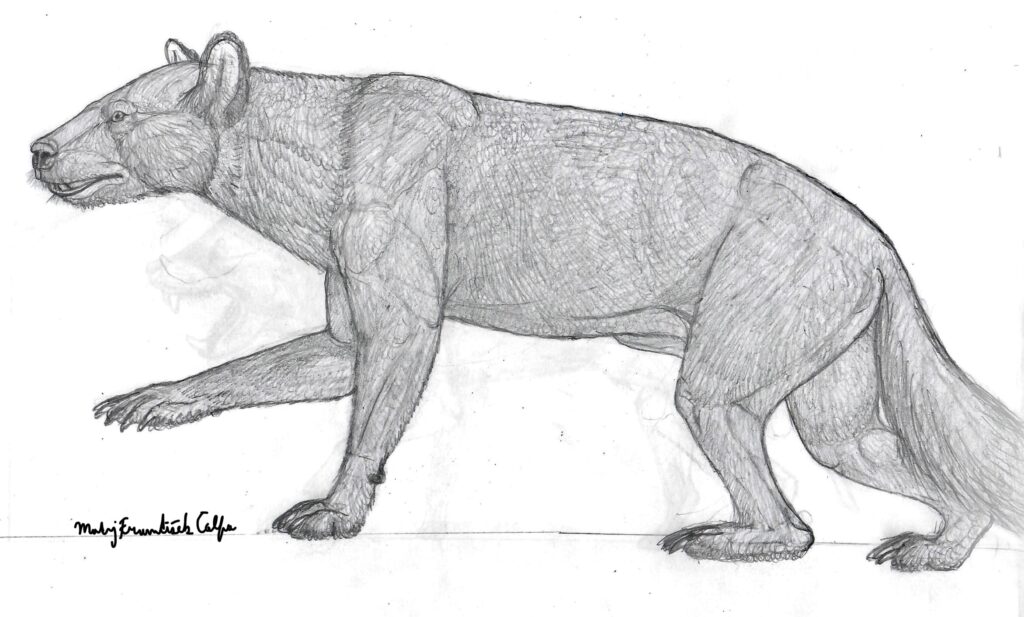
Here, a hypothetical skull is shown:
Right now, the question remains: Was Lonchocyon an amphicyonid or a true bear of the subfamily hemicyoninae? It’s not known yet. What is known however, is, that it represents an early offshoot of one of these groups, appearing quite early in their history (Late Eocene).
During the cenozoic, the suborder Caniformia has produced several lineages of carnivores, which cannot be compared to any living caniforms, hence, occupied a niche that either today remains vacant or is taken by a non-caniform. Such example would be the amphicyonids (bear-dogs) or the ursids (true bears) of the subfamily hemicyoninae. Contradictory to the name “bear-dog”, later, more specialized amphicyonids (amphicyoninae, thaumastocyoninae..) show a combination of skeletal traits and articulations rather resembling a mix between a bear and a big cat. Having a tendation to become hypercarnivorous, with the ability to deliver powerful, deep canine bite, combined with well developed pronation-supination movement, one would imagine these predators as solitary, ambush hunters and scavengers, much more like big cats and bears than any canid or hyaena. One representative of these robust carnivores is Ammitocyon:
Does this mean that this is exactly how we can imagine Lonchocyon to look like? Perhaps yes. Until more of its material is found and described it’s hard to tell with absolute certainty how exactly L. qiui was built.
This fascinating caniform remain to remind us of our humble knowledge of the history and evolution of the Caniformia clade, that in the past produced one of the most fearsome mammalian land predators that roamed the whole Afroeurasia and North America.
A medium dog-sized carnivore that is able to take down on its own ungulates, survives the harshest winters far north and does not hibernate..
Well, there is an actual animal like that, the wolverine (Gulo gulo).
Wild wolverine in Finland. Image: pixabay
The wolverine is a great example of an opportunistic predator and scavenger. It will feast on anything it stubles on, taking huge advantage of any opportunity given.. While most of the time it’s looking for smaller prey and carrion, it can take down ungulates much larger than itself such as reindeer. It’s also a scavenger, with a very acute sense of smell, capable of detecting carcasses covered up by snow with great accuracy.
With frost and water resistant fur and paws of very large size, that help them to stay on top of deep snow, its anatomy is greatly adjusted to the conditions of arctic and boreal forests.
Its lifestyle and habits are reflected in the skull and dentition. When compared to their closest relatives, it posseses enormously enlarged carnassials (P4 and m1) to crush bones and also, the upper molars are rotated in 90° angle which enables them to tear off frozen solid meat. Overall, the skull looks like that of an absolutely formidable predator.
The wolverine is a member of Mustelidae, that means that it’s basically a giant marten or a weasel rather than a small bear. The members of the weasel family are known to have absolutely enormous capabilities relatively for their body size, which is only confirmed in this species.
To look a little bit at its background and history, among mustelids, the wolverine is most closely related to martens (Martes) and fisher (Pekania). Together forming the subfamily Guloninae. To whose of these is the wolverine more closely related isn’t 100% sure. It’s likely that their common ancestor looked similar to the fisher Pekania occulta.
I had the idea of reconstructing some of its ancestors, or closely related extinct species. What came to my interest, is this fragment of the lower jaw of the ancestor of wolverine, being classified in its own species Gulo schlosseri, from Żabia cave in Poland.
To highlight the proportions and life appearance of this animal with the best possible accuracy, I combined this material with skeleton of modern wolverine, which gave me a good base for my drawing.
With a decent “strolling” pose and proportions, the muscles were added very roughly. I chose it yawning, to show its stunning hypercarnivorous dentition mentioned above.
Since the animal, in life would be all covered in fur, no muscular anatomy would be visible. This has made the job a bit easier. Here’s the outline of it.
Then, the details…
And finally, coloration. Althought color variations in individual wolverines can be suprisingly variable, for this one, I chose more darkly-toned individual. Perhaps, all these populations were darkly colored because of their shared ancestry with the fishers (Pekania)? This is a matter of speculation :).
Here we have it. The reconstruction of Gulo schlosseri, an ancestral wolverine from Early Pleistocene. Acting more or less like its living descendants, it surely would have been an amazing experience encountering one of these in their natural habitat.
Believe it or not, during the early Middle Miocene (here, I focus on roughly 16-14 million years ago, but overall it was for a longer period of time), large parts of Slovakia and Eastern Europe were covered by a massive sea called the Paratethys, with Carpathian mountains being just an archipelago within this massive inland water body!
It was in this setting, where numerous fossils of small-sized earless seals (Phocidae) were discovered. In the outskirts of Bratislava, near the peak of Devínska Kobyla, two species of one of these seals were described. Devinophoca claytoni and D. emryi. Together forming a now completely extinct subfamily Devinophocinae that possessed mixed characteristics with the other extant subfamilies of seals.
Here is shown my reconstruction of D. emryi. It was a seal of rather small size, being close to the Baikal seal. The missing parts of the skeleton and the general appearance is based on bunch of other extant seals such as Phoca, Pusa, Monachus and Neomonachus.
The reconstruction of Devinophoca emryi (skull lenght 11.95 cm)
Althought, it would look a lot like its modern counterparts, D. emryi possesed proportionally smaller skull and longer flippers. The latter might have been an unsuccessful evolutionary adaptadion that led to the eventual demise of this group. It has been suggested that Devinophoca might actually represent a very primitive relict of the common ancestors of other phocid seal species. That would mean that this is how we can imagine some of the first earless seals to look like. It lived in a tropical -subtropical environment of transitional shallow marine waters with coral-reef zones.
A sketch showing a pair of Devinophoca emryi in the Slovakian Central Paratethys.
Seals of the subfamily Phocinae can be divided into 5 exact groups based on morphology and ecology. (Representatives of other subfamilies can also be assigned, but its group can differ based on what criteria are used, as in Medditerrean monk seal (Monachus monachus)). Our seal, Devinophoca emryi, althought not belonging to Phocinae, based on morphology alone, falls exactly into the group 3. It’s reasonable to speculate that it was a bottom feeding seal, preying mostly on invertebrates such as mollusks, crabs, shrimp, amphipods and also fish.
The still-living Ribbon seal also falls into the ecomorphological group 3. Image: Michael Cameron
Devinophoca lived in a tropical-subtropical environment, similar to that of Hawaiian monk seal. Image: Kent Backman
Both species D. emryi and D. claytoni fall into different ecomorphological groups, therefore it is totally possible that they coexisted without any competition and niche overlap (for example hunting in different depths) as it’s also seen in some modern seals.
There is so much that could be additionally said, but for now, my time is up. Hoping to get back to this!
The skull in lateral and ventral view (scale bar 10 mm)
The orbital angle
Some time ago, in a forest near one village in Slovakia, I’ve found this skull. After looking at it for a bit, and drawing it, I suppose, that it most likely belongs to a dog, with a wolf-like head, perhaps a german shepherd, wolfdog, some northern breed or a mongrel of them… But not totally excluding the posibility of a wolf.
The lenght of the skull is a little smaller (21,1 cm) than that, what I found to be the average for the wolf as a species (23 – 28 cm), althought I’m not aware of the local wolves’ average sizes. The individual was already mature and probably not the youngest, since the teeth (P4, M1, M2) show large wear.
One characteristic commonly used to distinguish between wolves and the first domesticated dogs, is the orbital angle (shown in the picture above). The sharper the angle is the more “wolf-like” the head becomes. Some disagreement exists however, because apparently it’s a very variable feature, that must be taken with a grain of salt. I applied it to my skull, and it falls within the lower edge of a domesticated dog standards and higher edge of wolf’s.
As this is a very quick overview, and I’m an amateur on Canis skull morphology, feel free to refer to anything I missed out on!
I bet a lot of you are familiar with the Dire wolf (Aenocyon dirus). The biggest canine (member of caninae) ever, that roamed North america and preyed on the large megafauna of the Pleistocene Americas. Althought it wasn’t really closely related to the typical wolf, and hence it wouldn’t even look like one. What can be overlooked however, is that, during these times, the modern wolves (Canis lupus) not only existed in large numbers, but also were much more diverse, with very large and robustly built morphs extending as far north as the Beringia.
Beringia is usually devided to western (Russian side) and eastern (Alaskan side) part. Sea levels were much lower then, which meant that the bridge between the continents was connected. This article mostly focuses on the eastern part (Fairbanks area) but not only. During the late Pleistocene, (here focusing on the last 100,000 years) Beringia was nothing like it’s today. At that time, a mosaic of high productive steppe-tundra existed, which supported numerous species of herbs, grasses, lichens and even shrubs and trees… It was in this environment where a handful of megafaunal species could be found, from herbivores: mammoth (Mammuthus primigenius), bison (Bison priscus), caribou (Rangifer tarandus), woodland muskox (now considered Bootherium bombifrons), the wild yak (Bos mutus) and Yukon horse (Equus lambei) and on the western (siberian) side also the Lena horse (Equus lenensis) existed, among others. The guild of the largest carnivores was occupied by the now-extinct wolf ecomorph (Canis lupus), giant short-faced bear (Arctodus simus) Brown bear (Ursus arctos), cave and/or american lion (Panthera spelaea/atrox) and the scimitar cat (Homotheriumserum).
Two beringian wolf individuals with different color variations (typical grey-toned and lighter arctic form) with one of their prey items, the Yukon horse. Human sillhouette is ~1.8 meters tall.
Althought this environment was always more or less very cold, several climatic changes are recorded, with the Last Glacial Maximum (roughly somewhere between 26,000 to 18,000 years before present) being very prominent one. This means that overtime a lot of smaller changes in the faunal communities existed. From stable carbon and nitrogen isotope and radiocarbon analysis, the diets of species can be inferred. The beringian wolf was both an active predator and a scavenger, which enabled it to survive even the harshest periods. These wolf are considered to be horse/bison generalists, and competed for the same resources with other predators such as the scimitar cat (Homotherium serum). From the eastern Beringia (Fairbanks area), interestingly, during the whole Late Pleistocene only the wolf seemed to persisted the whole time, while for example felids (scimitar cat and lion) were only scarcely present when short-faced bears occupied the land (40,000 to 20,500 years before present). For comparison the latter seems to have been a caribou specialist, althought it is reasonable to assume that the specializations of different carnivores cannot be so stricly defined (if a predator was starving it would eat anything it stumbled upon).
The mammoth steppe was a very vast habitat in both area and time, which stretched through whole Eurasia and Alaska, so surely there must had been a lot of different local interactions and dynamics both between the predators themselves and their prey.
This environment produced, compared to its modern counterparts, an extreme form of the wolf (Canis lupus), that was adapted to scavenging and hunting on the large herbivores present. It tended to posses short, broad palates and larger carnassials relative to the skull size, altogether indicating adaptations typical for bonecraking behaviour, that would require the ability to produce large bite forces. Perhaps one of its good modern analogues is the Spotted hyaena of the african savannahs.
The beringian wolf
According to the genetic evidence the beringian wolves (even those found on the American side) were not related to the modern North American and Eurasian wolves, and most likely represent a lineage, now extinct, that once stretched through the whole Eurasia. For instance, several pre-LGM (Last Glacial Maximum) wolf specimens from Ukraine, Chzechia and Altai (Altai dog) region show genetic (mitochondrial DNA haplotype) similarities to some beringian wolves, which is opposed to the modern populations which show no relatedness.
Despite before-mentioned differences, in nature, it would look like a normal wolf. Here, two possible color variations are shown based on the environment (the standard grey-toned and a lighter arctic form). As one of their potential prey items, I drew the Yukon horse, which (based on one partial carcass) would be in life a small (~1.2 meters tall) horse closely resembling the tarpan and przewalski’s horse, perhaps with a thicker fur adapted to colder climates.
During the Early Pleistocene (roughly 1.3 million years ago), East Asia was home to a number of different large-bodied canines, including genera such as: Canis, Xenocyon, Cuon and Sinicuon. However, the exact taxonomic position of every specimen found is not completely clear…
Recently described partial skeleton (in 2012), with most of the vertebral column and rib cage missing, is an important one. It’s pretty obvious that this specimen is a member of the genus Canis, based on the material, it was assigned to Canis chihliensis, a species unique to China. While being much smaller in size than the modern wolf (Canis lupus), it also appears to be more primitive, thanks to less robust built of the limbs. The assumption of primitivness is based on the fact that canines (Canis, Cuon and Lycaon), in general tend to evolve to become more heavily built and adapted to hunting prey larger than themselves. Additionally, different specimen of C. chihliensis show signs of serious tooth wear and breakage as well as tibial fracture. All the trauma and infections seen in this canid are a good indicator for bone-crushing behaviour and living in packs (the specimen with a broken leg has a much higher chance of healing and surviving when it is cared for in a group), which is linked to hunting large prey.
This canid shared its habitat with numerous other ice age mammals, such as the hyaena Pachycrocutasp., pantherine cats, Nihewan wooly rhinoceros (Coelodonta nihowanensis), Steppe mammoth (Mammuthus trogontherii), the deer Eucladocerosboulei, horses and more…
Don’t know if you knew this, but the ecological niche of woodpeckers, that is, penetrating trees to reach and extract wood-boring invertebrates, has been also occupied by mammals. Living examples of such kind are: The marsupial genus of the-so-called trioks and Striped possum (Dactylopsila) and the lemuroid primate Aye-aye (Daubentonia). These species share several diagnostic traits that are associated with this kind of behaviour, such as: Massive, rodent-like incisors, used to rip off the bark of trees and one elongated, either third or fourth finger used to extract the insects out of the holes.
Interestingly, during the Paleogene (Paleocene to Oligocene) a whole order (and family) of mammals existed that specialized in such woodpecking behaviour, called Apatotheria. The members of Apatemyidae have been found all over the world. One species Heterohyus nanus, with different specimens being found in the Messel Pit even preserve soft tissue. This species (as well as others of its family) shows clear convergence between the formerly mentioned extant mammalian woodpeckers. They were small agile and active creatures spending much of their time foraging for food in the trees but being also able to run on the ground and leap short distances. Here are my attempts at reconstructing these critters:
Heterohyus nanus in lateral view (scale bar 5 cm)Heterohyus nanus in a somewhat more dynamic posture
One of the last known apatemyids, the genus Sinclairella, come from Arikareean North American Stage of the John Day Formation. When we compare its skull to the extant marsupial Dactylopsila (picture on the top), we find out that they look strikingly similar:
*Not to scale, http://digimorph.org/specimens/Dactylopsila_trivirgata , https://escholarship.org/content/qt15f034zz/qt15f034zz.pdf
The reconstruction of Sinclairella dakotensis
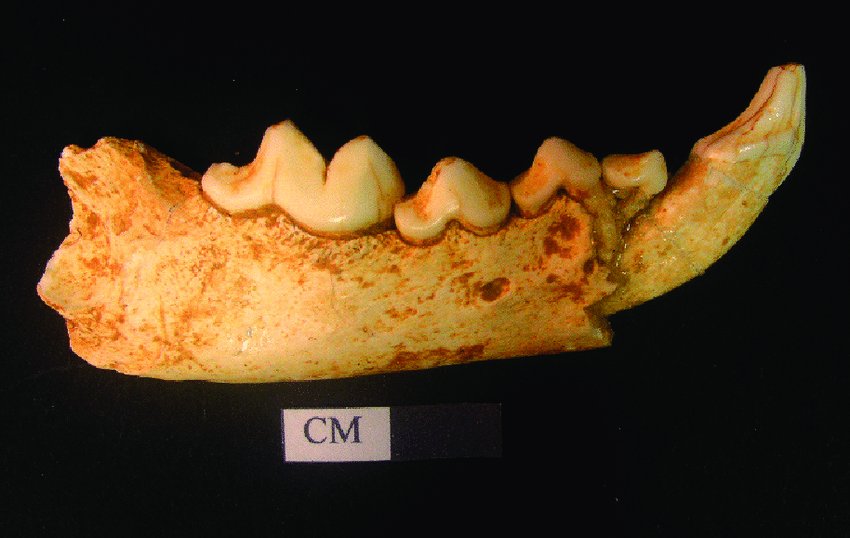
The two extant species of beaver are listed in the genus Castor. Historically this genus had a very wide distribution, in Eurasia ranging from Iberian Peninsula to China and in America from Alaska to northern Mexico.
However, the specimens of Castor from Late Miocene of China exhibit a distinct morphology that falls out of its norms. Therefore, a new genus has been created to list these specimens, called Sinocastor. Some differences between these genera includes: more elongated rostrum and neurocranium with the latter being not as broad, narrow postorbital region, (which are considered to be a more primitive traits) relatively longer incisive foramina and more…
If Sinocastor is a sister to Castor, it would meant that, the common ancestor of these two genera (therefore of a still-living beavers) may come from East Asia. Castor may have dispersed northeastward via Beringian Isthmus to North America and northwestward to Europe.
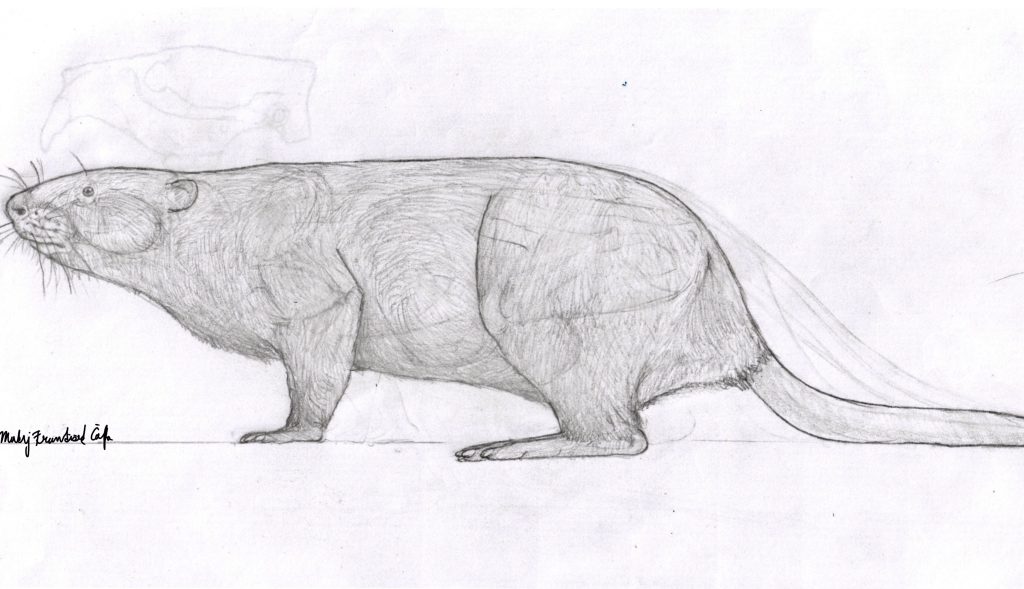
Here I show my reconstruction of Sinocastor with missing parts filled in by Castor fiber:

























{kind=link}
{kind=link}